![]()
Growth Control by Fat in Drosophila
|
||
|
|
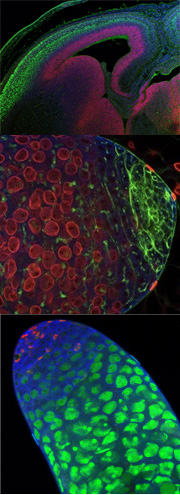
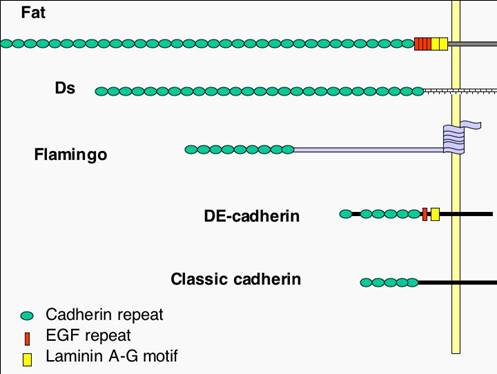
Growth control by the Drosophila tumour suppressor gene fat. Genetic studies have demonstrated fat is critical regulator of tissue growth in Drosophila, fat encodes a large transmembrane protein with 34 cadherin repeats, laminin G and EGF domains, and a poorly characterized cytoplasmic domain 11 (Figure 1). fat is called a Drosophila tumour suppressor gene because loss of fat leads to enormous overgrowth of larval tissues. We and others have recently found that loss of fat results in such dramatic overgrowths because it co-ordinately regulates both cell proliferation and apoptosis. An ortholog of Drosophila Fat, called Fat4, is well conserved to humans , suggesting our that understanding fat function in tissue growth in Drosophila is relevant to man.
The Hippo Pathway in DrosophilaThe Hippo kinase pathway is a conserved pathway that restrict tissue growth in flies and in man. It was discovered because, like fat, mutations in the Hippo pathway lead to massive overgrowth of Drosophila tissues . Expanded (Ex) and Merlin are related FERM-domain proteins that localize to apical junctions and semi-redundantly control cell proliferation by regulating Hippo activity. It is unclear how Ex and Merlin regulate Hippo activity. Hippo (Hpo), an MST-family kinase phosphorylates and activates Warts. Warts, an NDR-family kinase in turn phosphorylates and represses Yki. Yki is a transcriptional activator that regulates tissue growth by controlling the expression of Cyclin E, and the anti-apoptotic protein dIAP.
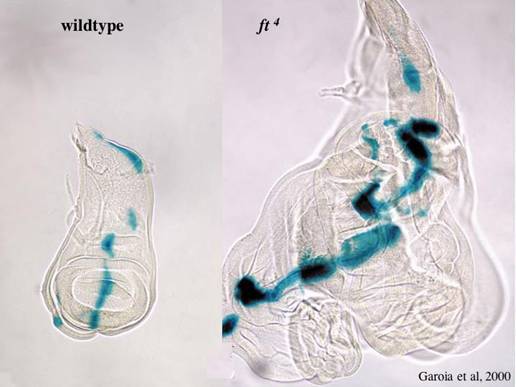
Until recently, Ex and Merlin were the most upstream known components of the Hpo pathway. Our recent studies indicate that fat controls tissue growth by regulating Hpo activity. We find that fat acts genetically upstream of ex in growth control, and regulates Ex localization at apical cell-cell junctions6. Ex is a large cytoplasmic protein that contains an N terminal FERM domain and three putative SH3 binding domains(Figure 1). Database analysis indicates that there is human homolog of Ex, FERM6, but it is currently uncharacterized. Like fat, loss of ex leads to tissue overgrowth in vivo in Drosophila. The vertebrate Hippo pathwayThe human orthologs of the Hpo kinase pathway components, hpo, and wts, can rescue their corresponding Drosophila mutants, suggesting that the Hippo pathway is conserved in mammalian cells . In support of this hypothesis, the human Hpo ortholog, MST2, can phosphorylate and activate the human Wts orthologs Lats1 and Lats2. Indications that this pathway may be tumor-suppressive in mammals include a tumor-predisposition phenotype (soft-tissue sarcomas and ovarian tumors) in mice lacking Lats1 , and suppression of transformation of NIH 3T3 cells by Lats2. Loss of Yap, the vertebrate ortholog of Yki, has recently been implicated in breast cancer and liver cancer. Thus understanding how the Hpo pathway is controlled may be important for understanding, and eventually treating, many human cancers. We have shown that Fat regulates the Hpo pathway and the membrane localization and stability of Ex . It is currently unknown how Fat regulates Ex localization, or how Ex regulates Hpo activity. Understanding how Fat regulates Ex localization is key to understanding how Fat and Ex regulate the Hpo pathway and is a current goal of our research. Fat (Ft) is very large, atypical cadherin that regulates planar polarity and growth in Drosophila. Loss of Ft leads to massive overgrowth of imaginal discs (see Figure 4). Loss of Ft also leads to planar polarity defects. Ft binds to another large atypical cadherin Dachsous (Ds) to regulate planar polarity.
Figure 3 Ft encodes an enormous cell adhesion molecule, of ~ 560kDa, which is a member of the cadherin family of cell-cell adhesion molecules.
|
|
|
|
Relevant Papers:
|
|
|
|
||
 |

.jpg)

.")

